
The Trouble with “Factors”
Craig Holdrege
From In Context #52 (Fall, 2024) | View Article as PDF
In the biological sciences many concepts are used habitually without careful reflection on the biases they inherently carry with them. One such concept is that of “factors.” In this article I want to consider “factors” as related to plant development and growth, with a focus on roots. What picture of organisms arises when we have analyzed their activities and relations into myriad factors? This consideration grew out of our ongoing research project on “Intelligence in Nature,” and I will discuss the implications of the factor-based thinking that colors current conceptions of plant intelligence.
Geotropism in Roots
“It can scarcely have escaped the notice of the most inattentive observer of vegetation, that in whatever position a seed is placed to germinate, its radicle [primary root] invariably makes an effort to descend towards the centre of the earth, whilst the elongated germen [shoot] takes precisely the opposite direction….” With these words, the English naturalist T. A. Knight introduces his 1806 letter to the Philosophical Transactions of the Royal Society of London, the premier scientific journal of his times. He reports on his observations and experiments concerning the way the plant relates to its environment during germination.
That the seed can be in any position when it germinates and the shoot then grows upward into the air and light, and the primary root grows downward into the soil, is a significant fact (even if noted by a most inattentive observer). If the tip of the root emerges out of the upper side of the seed, the growing root curves around so that the tip becomes pointed downward. If the plant is a species that forms a tap root (think of a dandelion or a carrot), the main root continues its downward growth.
Knight asks the question: Is this directional growth related to gravity? In asking this question he shows that he is not satisfied with the observation that primary roots usually grow toward the center of the earth. He knows something of Newtonian physics and has some idea about a gravitational force. Is this force working on the plant’s root? To answer this question he thinks of ingenious experiments in which he subjects germinating seeds to centrifugal force. Think of the experiences you had as a child on a fast, rotating merry-go-round and the feeling of being pulled away from the center of the merry-go-round — that’s centrifugal force. His idea was: If I subject the germinating seed to a centrifugal force, and subsequently the primary root no longer grows directly downward, I’ve shown — albeit indirectly — that normally the primary root grows down because of the earth’s gravitational force.
Based on this thinking, he constructed a number of different contraptions to test his hypothesis. In one experimental set-up he attached germinating bean seeds to the edge of a horizontal, rotating plate (imagine a record player). When the plate rotated rapidly with 250 revolutions per minute, he found that instead of growing straight downward roots grew outward and only slightly downwards. This confirmed to him that the roots were responding to the centrifugal force. This type of experiment has been repeated by many plant researchers and is a standard illustration in botany classes and textbooks. Knight goes down in the history of botany as the first person to have clearly demonstrated that plants are responding to gravity by growing vertically downward or upward. Later the terms geotropism and gravitropism were coined to explain the relation between plant growth and the earth’s center.
Other Tropisms
Any casual observer of roots also knows that by no means do they only grow down. Depending on the context, they can grow in all directions. So once you isolate in thought the idea of gravity as a factor influencing plant growth, and you are cognizant of at least some of the different ways roots (and stems) grow, you ask: What other factors could be involved? To find out, you need to do other kinds of experiments.
One particularly elegant and simple experiment was devised by Julius von Sachs (1832-1897), who is often considered the founder of modern plant physiology. Sachs knew, of course, that roots are highly dependent on water for their growth and the health of the whole plant. Can this be seen in the way they grow? He suspended a metal ring at an angle, covered the bottom with a fabric mesh, and filled what was now a container with moist saw dust (see illustration).
He placed peas (or other seeds) in the saw dust, kept the container in the dark, observed what the roots did. Ashis illustration shows, the roots grew first vertically down toward the center of the earth – demonstrating again geotropism. Then they penetrated through the fabric and for a very short distance continued to grow straight down. Very soon they began to curve to the side so that the roots came into contact with the moist underside of the mesh and grew along it. Sometimes they even curved upward and grew back through the mesh into the moist sawdust. (I repeated this experiment and made essentially the same observations.)
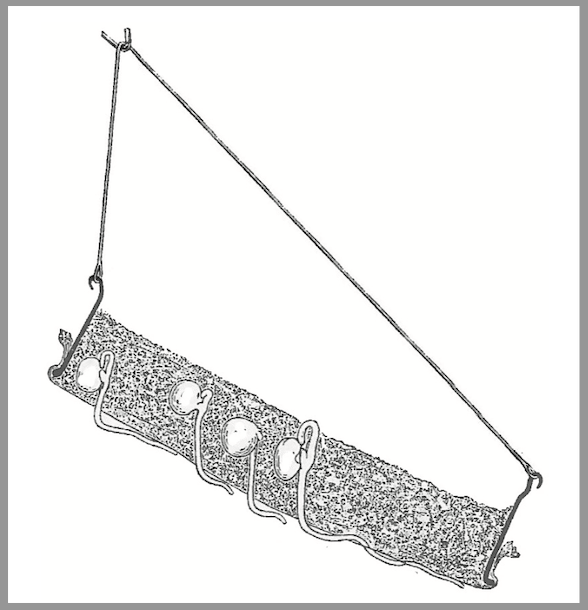
Julius von Sachs’ experiment; see text for explanation (from Sachs 1887, p. 715).
Here Sachs demonstrated that the growth of the roots relates to what he calls “contact stimulus” and moisture. If he kept the container in a space in which the air was saturated with water vapor, the roots continued to grow downward. Only when the air was less humid did they turn toward and grow along the mesh. So he concluded that hydrotropism (turning toward moisture) was dominant in this case, and contact stimulus (thigmotropism) secondary.
We now have geotropism, hydrotropism, and thigmo-tropism. It is easy to see that one can look in an ever more differentiated way at plant-environment relations. Here I’ll stay with roots and the soil environment: Do they actively grow away from light? What about soil pH (acidic or alkaline); compactness and texture (sand, silt, or clay); nutrients (phosphorous, nitrogen, potassium, etc.); organic matter; symbiotic relations (e.g. mycorrhiza); various organisms in the the soil?
Researchers have carried out numerous experiments to investigate these and many other aspects of the environment that the plant relates to through its roots. As the examples I just described indicate, in such laboratory experiments researchers attempt to create simplified conditions that allow them to focus on a particular aspect of the living organism and on a particular feature of its environment. In this process of investigation, they gain an ever more differentiated picture of the plant in its environment. The question that I’m interested in here is: How do researchers conceive of all the relations?
“Factors” and “Parts”
In his seminal and wonderfully detailed Lectures on the Physiology of Plants (1887), Julius von Sachs writes: “What I would like to emphatically lay down, is the fact that every phenomenon of life arises from two factors: on the one hand, from the structure transmitted from the mother organism, and, on the other, from external forces working on this structure” (p. 191). By means of the external influences, the “internal structure is impelled to the movements and performance of work specifically peculiar to it” (p. 190).
We can think of gravity, water, light, or soil nutrients as some of those external factors that work upon the plant. The plant, according to its structure, responds — in relation to gravity, roots grow down and shoots grow upward. It seems self-evident to think of what we call “environment” as external to the plant. The moment we have this conception we separate in thought environment from plant. The environment works on the plant and even impels the plant’s movements and performance — in Sach’s words. We think of a cause-and-effect relationship.
In this conceptual framework, the goal of scientific understanding is to discover and articulate the precise cause-and-effect relations between outer factors and inner conditions. Researchers analyze the organism ever more finely into its “parts” and the environment into specific “factors.” Ideally we could say: “Factor X causes the roots to do Y.” And then the task is to determine the causal chain of events within the organism through which the outer stimulus becomes translated into a specific physiological response. In contemporary biology, this is called discovering the underlying “mechanism.”
This causal way of thinking entails conceiving of “factors” as clearly circumscribed agents that affect the organism’s “parts” consistently in the same manner, eliciting the same response (effect) each time. This is the ideal of explanation that underlies modern biology, also in contemporary research on plant intelligence.
Botanist Angela Hodge describes in a review article many different experiments on plant responses to factors related to nutrients in the soil (Hodge 2009). She writes that “most of the information on root responses come from studies that have changed only one factor at a time and often use extreme contrasts.” In other words, the degree of disconnect from a plant growing in a natural context is substantial. Moreover, when researchers find a particular response in a particular species in a specific experimental setup, it doesn’t mean they will have the same results if they change the experimental conditions or consider a different plant species in the same kind of experiment. When Hodge surveyed the results of experiments related to a particular factor, she could not find general “rules” about how roots will relate to that particular factor. One can’t say factor X elicits response Y.
Here is a striking example: Arabidopsis thaliana, a small and inconspicuous species in the mustard family, is the ‘model’ plant with which thousands of researchers around the world experiment. More is known about Arabidopsis than about any other plant in terms of genetics and responses to the environment. Typically, researchers do experiments with one of a few main varieties of Arabidopsis. These varieties, generally called ecotypes, derive from plants taken from specific locations and environments, and they are morphologically and genetically distinct.1 They show the diversity within a plant species and its flexibility to change in relation to specific environments. When researchers work with one specific ecotype, it is easier to obtain repeatable results. The problem is that you don’t know whether what is valid for one ecotype is valid for another.
To get beyond this limitation, researchers in France carried out experiments with 73 different Arabidopsis ecotypes from many regions of the earth and different environments (Chevalier et al. 2003). They investigated how these ecotypes responded in experiments in which they were “starved” of the nutrient phosphorus. The plantlets were grown in petri dishes with artificial nutrient media — nothing resembling natural conditions. The focus was root growth. Under phosphorus-starved conditions half of the ecotypes exhibited less primary and lateral root growth; one quarter of the ecotypes responded in contrasting ways and only primary or lateral root growth was affected, not both; the remaining quarter did not show any response to phosphate starvation. There was no correlation between geographical region and response. Clearly, no general statement could be made in this lab experiment about the “factor phosphorus” on root growth in Arabidopsis.
Of course, no soil in nature has mineral nutrients in isolation from each other and all other aspects of the soil. So even if the researchers had obtained more consistent results, the question of their relevance to conditions in the natural environment would still be open.
The Problem of Isolating
In order to distinguish “factors” in a clear and distinct manner, we must isolate what becomes a factor from others in an experiment to determine whether the factor affects a specific aspect of plant growth and development. In the actual soil, however, we don’t find isolated factors, even though we may think about the soil as an agglomeration of factors.
In his seminal work, The Organism, Kurt Goldstein discusses how important it is for scientists to become keenly aware of the process and consequences of the experimental method. Why? Because in experiments with isolated phenomena, the results tell us about what is happening in that specific and narrow context. The experimental context is not revelatory of what is happening in the larger context from which it has been abstracted. Everything reveals what it is and does within a context; if the context changes, then other features appear. Goldstein writes:
By virtue of this isolating, dismembering procedure one can readily abstract and single out from living phenomena those phenomena on the physico-chemical “plane.” But the attempt to reintegrate the elements thus abstracted, to reorganize these split-off segments into the reality of living nature, is doomed to fail. This vain attempt, however, is made again and again, overlooking the fact that it is quite possible to understand the part on the basis of the whole, but that it is not possible to comprehend the whole on the basis of the parts. (1995, pp. 377-8)
Goldstein is asking us to confront an uncomfortable boundary or threshold in the attempts to understand things solely through analytical experimental science: We have the urge to study things in detail in order to gain a clear understanding of them and their relationships. To do so we focus on particulars that we have abstracted experimentally and in thought from some aspect of reality that interests us.
In this procedure we are no longer dealing with the reality that we originally wanted to understand, but some “part” or “factor” of that reality that is now appearing to us in a new and reduced context. The “factor” is an experimental phenomenon; we don’t know in what way it may be a member of the larger context from which it — as a part — arose. We run the danger of applying in an unaltered way the concepts we form in analytical experimentation to the larger whole.This is one of the big problems lurking in the background of claims such as, “scientists prove…”. You always have to ask the question: Under what specific conditions? The question that follows is: What does this have to do with a real-life situation?
Reification and Intelligence
Let’s look at the following description: “Plants are constantly bombarded with sensory inputs and receive numerous biotic and abiotic signals from their environment. Abiotic signals include gravity, light, water, temperature, oxygen, and carbon dioxide as well as other gases (Kiss 2007).” All such “signals” have been discovered through analysis and the process of isolation. These are then considered to be independent realities that together make up the plant’s environment that is external to it. The plant is pictured as a separate, independent entity that as such confronts its environment. The environment “bombards” the plant with substances and forces to which the plant responds. If it does so adequately, it thrives.
In the plant intelligence literature, this picture of plant-environment relation is given a certain kind of interpretation. A plant receives different signals — gravity is telling the roots to grow down, the pebble it hits tells it to grow around, a patch of higher moisture in a dry soil says “grow this way.” And there are hundreds more that a root has to deal with, not to mention all the internal signals that come from the above ground plant that are responses to the signals from the above ground environment. These signals can be highly conflicting. What is the plant to do in face of this bombardment? It must make decisions; it has to choose; it has to choose well if it is going to survive. The plant must be intelligent.
Tony Trewavas summarizes the prevalent view of plant intelligence: “Plant behavior is similar to cognition in an analogous sense to that of a human being. A plant continually gathers and updates diverse information about its environment, integrates this with information on its present internal state, and then makes decisions that reconcile its well-being with its environment” (2016).
The problem is that the plethora of inputs — “information” — that the plant is said to be gathering is a reification of the results of the processes of isolation and abstraction. The analysis of the environment into myriad factors and of the organism into separate parts and mechanisms is the work of the intellect. With the intellect we isolate and make clear and crisp distinctions. If we imagine all the products of isolation as independently real, then environment becomes for us an agglomeration of factors, and so does the organism. (Humpty Dumpty does not need to fall from the wall to become fractured. Unbeknownst to him, he is already pre-fractured into myriad parts long before he tumbles.) Given the assumption that separateness is fundamental and connectedness occurs secondarily, the manifest coherence, plasticity, and integrative nature of the organism-environment relation becomes an ever greater riddle.
Current plant intelligence proponents deal with the task by imputing human capacities into plants. Plants become human intellects in the guise of plants. Plant physiologists who want to stay in the tradition of mechanistic science and reject plant intelligence arguments are generally satisfied with discovering ever more details about the workings of the plant in relation to the environment. They continue to find new and interesting phenomena and can outline mechanisms that reveal themselves in limited experimental conditions. Yet, as the example with Arabidopsis ecotypes shows, when researchers expand research by taking different types of organisms into account or vary the experimental conditions, new phenomena arise that call into question any straightforward cause-effect “explanations.”
From Separateness to Connectedness
It is something very real when we human beings feel separate or disconnected from the world. Much is a riddle to us. We ask questions. We search for answers. Before we act, we are often uncertain, we ponder and weigh different possibilities of action. We are not simply embedded in a world in which all is clear. We try to overcome in one way or another the gap or gulf we sense between ourselves and the world. It is one thing to sense our predicament as human beings and take it seriously. It is another thing to treat our felt separateness and alienation as something fundamental in the world and then, explicitly or implicitly, seek to instantiate it in our science.
There is certainly justification for conceiving of physical bodies as being separate things. We can demarcate the boundaries of rocks, chairs, and human bodies. We can easily consider all the discrete things in the world — individual bodies — as separate from each other. As a body I can run into other bodies. I don’t merge with the chair I’m sitting on. No doubt. If I model my way of thinking about the world on this character of experience, then I conceive of discrete bodies interacting. That’s how I try to make sense of the world. This is a prevailing habit of thought that gives us “factors” and “parts.” It goes much further when in school or through popular media we are led to conceive of atoms (incorrectly from the standpoint of modern physics) as discrete physical particles, or genes (incorrectly from the standpoint of current molecular biology) as discrete entities populating a genome. This thought habit informs how we conceive of cause-effect relations in narrow terms — one circumscribed entity affecting another circumscribed entity. It is the basis of the conception that reality is built from the bottom up. There may not be turtles all the way down but there are entities all the way down. It follows that explanations must show how a given phenomenon is caused by the interaction of its parts.
But let’s think again about bodies — the chair, the rock, my body. All these bodies exist in relation to the totality of the earthly environment. They are bodies in gravity and rest on the earth. They consist of the “stuff” of the earth. They have a context. To conceive of bodies as separate is to isolate one important feature of their existence and to then model the way we think about them according to this feature. We can, however, from the outset, think in terms of context and relatedness that any given “thing,” any phenomenon, reveals. Nothing compels us to give preference to separateness. Take the water content of our body. Our “solid” body consists of about 60% water. This is not a static content. We are continually taking up water out of the environment and giving off water, as fluid or water vapor, to the environment. We are embedded, even though we’re not fish, in a water world.
This is also the case with air that we inhale and exhale; we are part of the earth’s atmosphere. And we are wholly embedded in the warmth environment of the earth. I hold a cool stone and it warms while my hand cools. My body generates heat and gives it off to the environment. As bodies we belong to the larger earth. With these few examples I want to direct attention to the reality of connectedness, of belonging. It is easy to say that everything is connected. But there is a real challenge here. If we don’t want to remain vague or be satisfied with general impressions, we delve into the world through observation and thinking. The more we do this the more we encounter differentiation and distinction. How do we move in this rich world without creating artificial boundaries? In biology the ideas of factors, mechanisms, and material cause-and- effect relations are convenient frameworks to think in. And from the point of view of manipulation, they prove highly efficacious. But they are also limiting inasmuch as our concern is deeper and comprehensive understanding.
How do we move beyond these limitations? If we don’t think in these terms, what do we do? This is the riddle that presents itself when we intimate that connectedness is fundamental in the world. What does connectedness-inquiry look like? For me this is an ongoing task of trying, of forming faculties that are nascent, and seeing where it all leads. I have little doubt that this undertaking, even in its beginnings, opens up fresh insights into the nature of organismic life. My colleagues and I hope to show in subsequent essays how it leads to more differentiated conceptions of intelligence in nature.
Notes
1. Ecotypes are known as “accessions” in the Arabidopsis literature.
References
Chevalier, F. et al. (2003). “Effects of Phosphate Availability on the Root System Architecture: Large-scale Analysis of the Natural Variation between Arabidopsis Accessions.” Plant, Cell and Environment, vol. 26, pp. 1839-50.
Goldstein, Kurt. (1995) The Organism. New York: Zone Books. (The original German book was published in 1934 and the first English version in 1939.)
Hodge, Angela. (2009). “Root Decisions”. Plant, Cell and Environment, vol. 32, pp. 628-40.
Kiss, J. Z. (2007). “Where’s the Water? Hydrotropism in Plants.” Proceedings of the National Academy of Sciences, vol. 104(11), 4247-4248.
Knight, T. A.(1806). “On the Direction of the Radicle and Germen during the Vegetation of Seeds.” Philosophical Transactions of the Royal Society of London, January 9, pp. 99-108.
Sachs, Julius von. (1887). “Lectures on the Physiology of Plants.” Oxford: Clarendon Press. (Facsimile reprint 2018 by Forgotten Books; forgottenbooks.com)
Trewavas, T. (2016). “Plant Intelligence: An Overview.” BioScience vol. 66, pp. 542-51. doi:10.1093/biosci/biw048