
The Giraffe’s Short Neck
Why Evolutionary Thought Needs a Holistic Foundation
Craig Holdrege
From In Context #10 (Fall, 2003) | View article as PDF
This essay is part of a larger monograph on the holistic biology of the giraffe, which appears in our Nature Institute Perspectives series and in Craig Holdrege’s book, Seeing the Animal Whole — And Why It Matters, published by Lindisfarne Books in 2021.

Figure 1. Giraffe in a “classic” feeding position, extending its neck, head, and tongue to reach the leaves of an Acacia tree. (Tsavo National Park, Kenya; drawing by C. Holdrege)
View some of our other articles on whole-organism biology
Lamarck and Darwin
Once scientists began thinking about animals in terms of evolution, the giraffe became a welcome — and seemingly straightforward — example. It is as if the giraffe’s long neck was begging to be explained by evolutionary theorists.
One of the first evolutionary thinkers, Jean-Baptist Lamarck, offered a short description of how the giraffe evolved in his major work, Philosophie Zoologique, which was published in 1809:
It is interesting to observe the result of habit in the peculiar shape and size of the giraffe: this animal, the tallest of the mammals, is known to live in the interior of Africa in places where the soil is nearly always arid and barren, so that it is obliged to browse on the leaves of trees and to make constant efforts to reach them. From this habit long maintained in all its race, it has resulted that the animal’s forelegs have become longer than its hind-legs, and that its neck is lengthened to such a degree that the giraffe, without standing up on its hind-legs, attains a height of six meters. (Quoted in Gould 2002, p. 188)
In Lamarck’s view, we must imagine a situation in the past where the best food for browsing mammals was higher up in trees, the lower vegetation having been eaten by other animals. The ancestors of the giraffe — which we should imagine like antelopes or deer — needed to adapt their behavior to this changing environment. As Lamarck wrote, “variations in the environment induce changes in the needs, habits and modes of life of living beings ... these changes give rise to modifications or developments in their organs and the shape of their parts” (p. 179). So Lamarck imagined that over generations the habit of continually reaching for the higher browse produced in the giraffe’s ancestors a lengthening of the legs and neck.
A little over 60 years later, Charles Darwin commented on giraffe evolution in the sixth edition (1872) of his seminal book, Origin of Species:
The giraffe, by its lofty stature, much elongated neck, forelegs, head and tongue, has its whole frame beautifully adapted for browsing on the higher branches of trees. It can thus obtain food beyond the reach of the other Ungulata or hoofed animals inhabiting the same country; and this must be a great advantage to it during dearths.... So under nature with the nascent giraffe the individuals which were the highest browsers, and were able during dearth to reach even an inch or two above the others, will often have been preserved; for they will have roamed over the whole country in search of food.... Those individuals which had some one part or several parts of their bodies rather more elongated than usual, would generally have survived. These will have intercrossed and left offspring, either inheriting the same bodily peculiarities, or with a tendency to vary again in the same manner; whilst the individuals, less favoured in the same respects will have been the most liable to perish.... By this process long-continued, which exactly corresponds with what I have called unconscious selection by man, combined no doubt in a most important manner with the inherited effects of the increased use of parts, it seems to me almost certain that an ordinary hoofed quadruped might be converted into a giraffe. (Darwin 1872, pp. 177ff.)
In many respects this is a classic formulation of how Darwin viewed evolution: every species consists of individuals that show considerable variations. Under certain environmental conditions particular variations will be most advantageous. Natural selection weeds out the unadapted and the best-adapted survive. These variations become dominant in the species and so it evolves. In the case of giraffes, times of drought and arid conditions give an advantage to those animals that can out-compete others by reaching the higher, untouched leaves. They form the ancestral stock of the animals that evolve into giraffes.
Interestingly, Darwin believed in the “inherited effects of the increased use of parts” — a very “Larmarckian” view. Lamarck argued for the inheritance of acquired characteristics. Darwin felt that this was key to explain giraffe evolution; otherwise there is no guarantee that longer features in one generation will have an effect on subsequent ones. But this view of the inheritance of acquired characteristics is rejected by mainstream Darwinists today.
The Long Neck as a Feeding Strategy
The idea that the giraffe got its long neck due to food shortages in the lower reaches of trees seems almost self-evident. The giraffe is taller than all other mammals, can feed where almost no others can, and therefore has a distinct advantage. It seems compelling to say that the long neck and legs developed in relation to this advantage. Why else would the giraffe be so tall? You find this view presented in children’s books, in web descriptions of the giraffe, and in textbooks.
But just because this explanation is widespread does not mean it is true. In fact, this “self-evident” explanation retains its ability to convince only as long as we do not get too involved in the actual biological and ecological details. Various scientists have noticed that this elegant picture of giraffe evolution dissolves under closer scrutiny. Here are a few examples of my and their objections:
1) Since the taller, longer-necked, evolving giraffe ancestors were also larger and heavier, they would need more food than the animals they’re competing with. Wouldn’t this counterbalance their advantage in times of dearth? Would they really have any advantage over smaller members of the same and other species? Moreover, it is absurd to assume that only the leaves on high branches were available to the giraffe during a drought. Had this been the case, then the multitude of browsing and grazing antelope species in Africa would all have gone extinct (or never evolved in the first place). So, even without growing taller, the giraffe ancestor could have competed on even terms for those lower leaves.
2) Male giraffes today are up to one meter taller than female giraffes; newborn and young giraffes are much smaller. The moment this sexual dimorphism manifested in the evolution of the giraffe, it would have been the males that could have reached the higher branches. The females and young animals would have died and the species would have gone extinct (Pincher 1949).

Figure 2. Giraffe feeding at about shoulder height — the most prevalent height at which giraffes feed. (South of Moremi Game Reserve, Botswana; drawing by C. Holdrege.)
3) If giraffes evolved by eating high foliage during times of drought and maximal competition for food, one would expect that giraffes today would also feed from the high foliage during these times in order to avoid competition. Males usually feed at greater heights than females and the results of one study show a surprising spread (Ginnett and Demment 1997). Male giraffes fed nearly half of the time at heights of almost five meters, that is, in the “classical” long-necked giraffe posture. In stark contrast, females fed around seventy percent of the time at belly height or below, which the theory demands they should not be doing. These researchers did not report on the seasons in which they made these observations, so their results are of little help in discerning whether, for example, males feed at greater heights mainly during droughts.
A variety of other studies show that giraffe feeding habits vary according to place and time (reviewed in Simmons and Scheepers 1996). Giraffes move seasonally, and in the dry season in East Africa they tend to seek out lower valley bottoms and riverine woodlands. There they usually feed from bushes at or below shoulder height (about two and one half meters in females and three meters in males). Fifty percent of the time they fed at a height of two meters or less, which overlaps with the feeding zone of larger herbivores such as the gerenuk and the kudu (Leuthold and Leuthold 1972; Pellew 1984). During the rainy season, when there is abundant browse at all levels, giraffes are more likely to feed from the higher branches, browsing fresh, protein-rich leaves. Other studies also show that giraffes do most of their feeding at about shoulder height, with their necks positioned nearly horizontally (Young and Isbell 1991; Woolnough and du Toit 2001; see Figure 2). So it looks as though giraffes are not using their long necks the way the theory demands. And they use them even less to reach heights in the dry season, when the theory demands they should need them most!

Figure 3. A goat does not require a long neck to feed on twigs and leaves of an oak tree. (Drawing by C. Holdrege after a photo in Butzer 2000.)
4) There are other ways to reach the high foliage of trees. Goats, for example, are known to climb into trees and eat foliage (see Figure 3). Why didn’t tree-climbing leaf-eaters (folivores) develop in the savannah? They would have had the advantage of feeding at all levels easily and been in that respect more adaptable than the highly specialized giraffe. The long-necked gerenuk, an antelope, often stands on its hind limbs and browses, reaching heights of two meters and more. The much larger and heavier elephant even stands sometimes on its back legs and extends its trunk to reach high limbs — but no one thinks that the elephant developed its trunk as a result of selection pressures to reach higher food.
In sum, there is nothing in this theory that shows a compelling link between leg and neck lengthening and feeding on high limbs. Just because giraffes have long necks and long legs and can reach food high in the trees does not mean that a need to reach high browse was a causative factor in the evolution of those characteristics.
Clearly, both Darwin’s and Lamarck’s conceptions of giraffe evolution were highly speculative. The idea that giraffes developed longer legs and necks to reach higher food seems plausible, even compelling, as long as we do not (1) think the idea through in all its implications and (2) take into account essential observations of giraffe behavior and ecology. In the end, the idea is neither logically compelling nor based on fact.
Alternative Explanatory Attempts
Pincher (1949), after critiquing Darwin’s explanation, suggests that the “most extraordinary feature of the giraffe is not the length of the neck but the length of the forelegs.” By developing long legs, the giraffe has acquired a huge stride so that it can move relatively fast for its size. This has left the giraffe with only one predator — the lion. Pincher therefore explains the “excessive length of its fore-legs as the effect of natural selection acting continually through the hunter-hunted relationship, as in the case of hoofed mammals generally.” The neck, in turn, followed the lengthening legs so that the giraffe could still reach the ground and drink.
It is strange that Pincher is able to critique Darwin’s view so clearly and yet doesn’t recognize that he is proposing the same type of inadequate explanation. The giraffe ancestor could just as well have developed greater bulk or more running muscles, both of which would have aided in avoiding predators. The fact is that despite its size and long stride, the giraffe is still preyed upon by lions. And as one study of one hundred giraffes killed by lions in South Africa showed, almost twice as many bulls were killed as cows (Pienaar 1969; cited in Simmons and Scheepers 1996). The longer stride of bulls evidently doesn’t help them avoid lions better than the shorter legged females. Who knows whether their long stride may in some way make them more vulnerable? Another speculative idea into the wastebasket.
Brownlee (1963) speculates that the lengthening of the limbs and neck in the giraffe give the giraffe a relatively large surface area, which should allow it to dissipate heat. This would be of advantage in the hot tropical climate, so that the tendency toward lengthening would have been encouraged by natural selection, since the largest animals would have been best able to survive heat waves.
As in the other suggested “explanations,” the central question is, Is Brownlee’s idea rooted in reality? Because of its long legs and neck, the giraffe appears to have a large surface area. But surface area alone is not important; it is the relation of the heat producing volume to surface area that is crucial. A small animal has a small volume in relation to a very large surface area, while a large animal has a very large volume in relation to its relatively small surface area.* Now the giraffe is a very large animal with a barrel-shaped torso. Although its neck is long, it is also voluminous; only the lower parts of the legs, which carry relatively few blood vessels, would act to enlarge the surface-to-volume ratio substantially. Krumbiegel (1971) estimates that the ratio of volume to surface in the giraffe is 11:1, compared, say, to a smaller, long-necked antelope, the gerenuk, which has a ratio of 4.7:1 (similar to the human). In other words, despite appearances, the giraffe still has a very large volume in relation to its surface area and its unique form provides no grounds to think that it evolved in relation to dissipating heat.
More recently, Simmons and Scheepers (1996) proposed that sexual selection has caused the lengthening and enlarging of the neck in males. These scientists place their ideas in relation to known facts and point out shortcomings in relation to larger contexts — a happy contrast to the other hypotheses we’ve discussed. They describe how male giraffes fight by clubbing opponents with their large, massive heads; the neck plays the role of a muscular handle. The largest (longest-necked) males are dominant among other male giraffes and mate more frequently. Since long-necked males mate more frequently, selection works in favor of long necks. This would also help explain why males have not only absolutely longer, but proportionately heavier heads than females. This hypothesis seems consistent with the difference between male and female giraffes. At least it gives a picture of how the longer neck of males can be maintained in evolution. But it doesn’t tell us anything about the origin of neck lengthening in giraffes per se — the neck has to reach a length of one or two meters to be used as a weapon for clubbing. How did it get that long in the first place? Moreover, the female giraffe is left out of the explanation, and Simmons and Scheepers can only speculate that female neck lengthening somehow followed that of males. In the end, the authors admit that neck lengthening could have had other causes and that head clubbing is a consequence of a long neck and not a cause.
Does the Giraffe Really Have a Long Neck?
All the above explanations of the evolution of the giraffe’s long legs and long neck are unsatisfying. Each of the authors sees problems in other explanations, but remains within the same explanatory framework when putting forward his own hypothesis. No one sees the necessity for stepping outside the framework and looking at the difficulties of the overall approach. The scientists abstract individual features (long neck, long legs, large surface area) and consider them in isolation from the rest of the organism. The individual feature is then placed into relation to one purported causal factor in the environment (drought, heat, predator avoidance, male competition). The link of individual feature to environmental factor is supposed to explain the evolution of that feature.
But this is a highly problematic procedure. The giraffe’s neck carries out a variety of functions — it allows feeding from high branches, serves as a weapon in males, brings the head to elevated heights that give the giraffe a large field of view, is used as a pendulum while galloping, and so on. Virtually all structures and organs in the animal body are multifunctional and interact dynamically with other multifunctional structures and organs. When scientists pick out a single function and focus solely on it to explain a multifunctional organ, their explanation can only be inadequate. This is comparable to believing you can paint a richly-nuanced, colorful rendition of a landscape with one color. It just does not work.
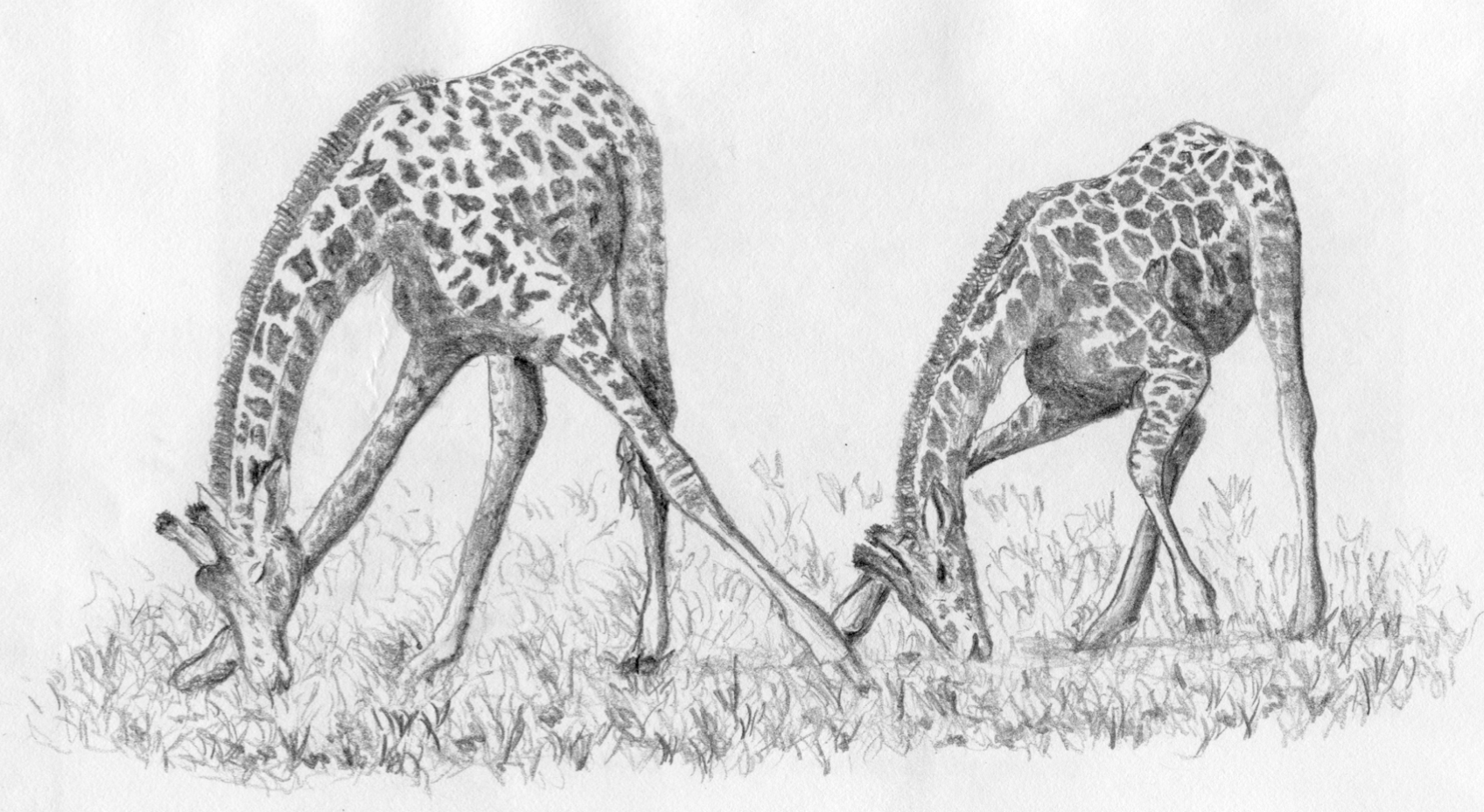
Figure 4. “Short-necked” giraffes grazing. Giraffes can only reach the ground with their mouths to drink or graze by splaying their front legs (left) or splaying and bending their legs (right). (Drawing by C. Holdrege after a photo in Dagg and Foster 1982.)
I sometimes wonder why no one has maintained that the giraffe has, in reality, a short neck. If you observe a giraffe drinking or, as they occasionally do, grazing close to the ground, then you know what I mean (see Figure 4). Giraffes do not drink often, but when they do, they have to either splay their forelegs to the side or bend their forelegs strongly at the wrist joint. Both procedures take time and are awkward for the giraffe. But only in this way can it get the tip of its mouth down to the surface of the water. So, looked at from the perspective of drinking, the giraffe has a very short neck. Antelopes and zebras reach the ground without bending their legs, and the long-legged elephant has its trunk to compensate for its short neck. Only the giraffe (and its rain forest relative, the Okapi) have necks that are so short relative to their legs and chest that they must splay or bend their legs.
So why hasn’t the giraffe become famous for its manifestly short neck? Why don’t we have evolutionary hypotheses explaining how the giraffe got its short neck? It is because the giraffe’s neck, in other respects or from other perspectives, is long. No other mammal has such a long neck in absolute terms or in relation to the length of its torso. We all have seen (in life or in pictures) and been amazed by the standing giraffe, its long neck sailing skyward, in comparison to which the ungainly, short-necked drinking giraffe appears as exceptional, almost unfortunate behavior.
Whether the neck is long or short depends on our perspective and on the behavioral or anatomical context we are focusing on. We only understand the giraffe when we view it from various perspectives and let the giraffe show different aspects of its being. The moment we focus solely on the “long neck” — and on it solely in terms of a food-gathering or some other strategy — we’ve lost the reality of the giraffe.
Reality is richer than such explanations. The explanation may be coherent and logical, but what it explains is not the thing itself but a specter of it — the isolated aspect that has been abstracted from the whole organism. In reality, the organism as a whole evolves; all its parts are multifunctional, facilitating its interactions with its complex, changing environment. If we don’t consider all partial aspects within this larger context, we can only have inadequate explanations void of life.
In sum: the whole project of explaining the evolution of an animal by abstracting from the whole leads to unsatisfying, speculative ideas on the one hand, and to conceptual dissolution of the unity of the organism on the other. A more adequate understanding requires that we first investigate the organism as a whole and how its members interrelate and interact within the context of the whole organism and its environment. This holistic understanding can then form the starting point for thinking about the evolution of the animal. The evolutionary biologist Dobzhansky’s famous statement that “nothing in biology can be understood except in light of evolution” is a grand claim, which I believe is, in the end, true. But we have a lot of work to do before we get there, and we should not be satisfied with short-cut evolutionary “explanations.” Another consequence of the usual way of explaining is that the organism itself is atomized into individual characteristics, each having its own explanation. Each part takes on a quasi-reality of its own, while the whole organism — which brings forth and gives coherence to the parts — degenerates into a kind of epiphenomenon, a mere composite of the surviving parts that “really” count.
If evolutionary thought is to have a solid foundation, we must establish this firm grounding in holistic understanding. As it is, stories of the evolution of traits seem compel-ling until you look for their context and foundation in the world and discover a pool of quicksand. As Simmons and Scheepers remark about Darwin's idea of giraffe evolution, “it may be no more than a tall story.”
* Assuming for the sake of explanation a spherical body, the two-dimensional surface grows as a function of the square of the radius, while the volume — being three-dimensional — grows as a function of the cube of the radius. A small sphere with a radius of 2.5 cm (1 inch) has a volume-to-surface ratio of 0.8:1. A much larger sphere with a radius of 50 cm (about 20 inches) has a volume-to-surface ration of 16.7:1.

For more of Craig’s work on the giraffe, see the article “The Giraffe in Its World” and the monograph “The Giraffe’s Long Neck.” Other articles on whole organism biology can be viewed here, or you can find Craig’s latest book on the topic, Seeing the Animal Whole—And Why It Matters, at our bookstore.
References
Brownlee, A. (1963). "Evolution of the Giraffe," Nature vol. 200, p. 1022.
Butzer, Karl (2000). "The Human Role in Environmental History," Nature vol. 287, pp. 2427-2428.
Dagg, Ann Innis, and J. Bristol Foster (1982). The Giraffe: Its Biology, Behavior and Ecology. Malabar FL: Krieger Publishing Company.
Darwin, Charles (1872). Origin of Species. Sixth Edition.
Ginnett, Tim, and Montague Demment (1997). “Sex Differences in Giraffe Foraging Behavior at Two Spatial Scales,” Oecologia vol. 110, pp. 291-300.
Gould, Stephan Jay (2002). The Structure of Evolutionary Theory. Cambridge, MA: Belknap Press.
Krumbiegel, Ingo (1971). Die Giraffe. Wittenberg (Germany): A. Ziemsen Verlag.
Leuthold, Barbara, and Walter Leuthold (1972). “Food Habits of Giraffe in Tsavo National Park, Kenya,” E. Afr. Wildl. J.vol. 10, pp. 129-141.
Pellew, Robin (1984). “The Feeding Ecology of a Selective Browser, the Giraffe (Giraffa camelopardalis tippelskirchi),” J. Zool., London vol. 202, pp. 57-81.
Pincher, Chapman (1949). “Evolution of the Giraffe,” Nature vol. 164, pp. 29-30.
Simmons, Robert, and Lue Scheepers (1996). “Winning by a Neck: Sexual Selection in the Evolution of the Giraffe,” The American Naturalist vol. 148, pp. 771-786.
Woolnough, A. P. , and J. T. duToit (2001). “Vertical Zonation of Browse Quality in Tree Canopies Exposed to a Size-Structured Guild of African Browsing Ungulates,” Oecologia vol. 129, pp. 585-590.
Young, Truman, and Lynnsebell (1991). “Sex Differences in Giraffe Feeding Ecology: Energetic and Social Constraints,” Ethology vol. 87, pp. 79-89.