
The Forming Tree
Craig Holdrege
From In Context #14 (Fall, 2005) | View article as PDF
The trees you see below are in their winter habit. Without leaves, the form of the crown displays itself through the intricate branching pattern of the limbs. Each of these trees has a history behind it and the crown form reveals some of that history. But the history is no straightforward matter. It has different facets and in each particular tree is unique.
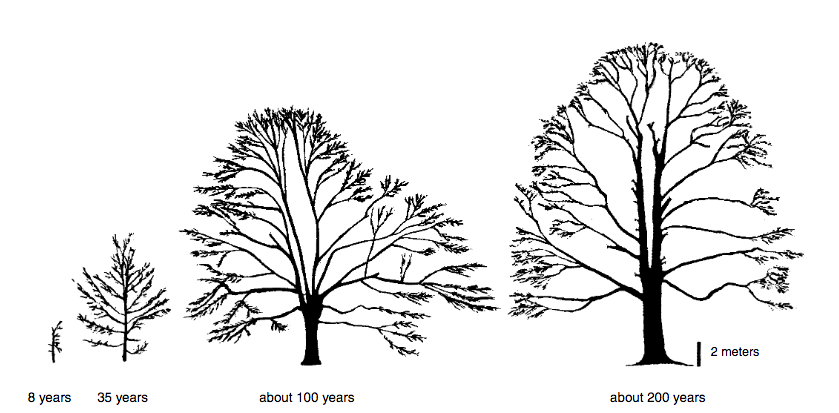
Figure 1. Schematic depiction of the growth of an individual tree, a European beech (Fagus sylvatica). (After Gleissner 2005, p. 66)
First, each tree belongs to a species. As a red oak or a white ash, a tree is part of a specific hereditary current that connects it with all other members of the species. Although a species has considerable plasticity and shows an often surprising variety of forms, it is nonetheless usually possible, with a bit of practice, to identify a tree species in the winter through its bark, branching pattern, buds, and so on.
The particular shape of the crown and the size of the trunk relative to the crown in an individual tree express a different facet of the tree’s history. A tree’s crown develops over time and no broad-leafed tree maintains the same shape when it grows from a sapling to a 20- or 100-year-old tree (see Figure 1). While growing, the shape transforms. All the trees you see in the figure on the opposite page had, as young trees, branches growing out of the trunk near to the ground. But all of these branches have since died off. As the trunk grew in diameter, the bark grew around the scar where the branch had separated from the trunk. The branchless lower stretches of the old trunk therefore no longer reveal outwardly the tree’s growth history. The tracks are present, however, as knots deeper within the wood.

Figure 2. A variety of tree forms. (Sketches by C. Holdrege)
The trees in Figure 2 vary greatly in shape, and the crown of some trees is markedly asymmetrical. You might even call them misshapen. To understand these forms you have to look not only at the growth process of the individual tree, but also at its growth in relation to the environment. Figure 3 provides a partial solution to the riddle of these enigmatic forms — you no longer see each tree by itself but within a group of trees. Each tree can in reality only be understood when you see it as part of a larger whole. Viewing the tree forms in isolation (Figure 2), you recognize that something isn’t quite right, but then, seeing them in context, you realize — with an element of surprise, relief and a sense of resolution — that everything is right. The individual trees fit together and form, as a group, one large crown.

Figure 3. The five trees from Figure 2 depicted in context — as tree groups in which the trees together form a crown. Left: white ash, American elm, and pignut hickory; right: red oak and white oak. (Sketches by C. Holdrege)
Evidently, trees growing up in close proximity relate to one another. A tree does not have a predestined shape that it has to achieve. Rather, it develops in relation to a specific constellation of organisms and qualities (light, water, soil, exposure) constituting its environment. It is a remarkable phenomenon that different tree species can grow in concert to form an overriding crown of which each is a part. When you study tree growth, you recognize that this co-development occurs largely in relation to light.
All plant growth is connected with light. In trees, the trunk brings the tree up into the light-filled atmosphere. But, as a rule, a tree trunk grows straight up; it is not directed toward the sun as a source of light, which in fact moves daily across the sky. This growth straight upward is known as negative geotropism, since the trunk grows directly away from the center of the earth. The blossoms of numerous wildflowers, in contrast, follow the path of the sun during the day; they exemplify positive phototropism — growing toward the light source.
As the tree trunk grows straight upward, it sends off side branches and, eventually, in most broad-leafed trees, the main trunk itself divides into smaller branches (unlike conifers such as spruce, fir, and hemlock, which usually maintain a central vertical trunk throughout their lives). Through this ongoing upward growth and branching, the tree form arises. Exactly how it arises depends on the particular context. (For more examples of how trees forms within the context of their habitat see Buess 1992.)
Figure 4 depicts two white oaks with dramatically different forms. The broad-crowned oak grew as a free-standing tree at the edge of a pasture. It had, as a young tree, no neighbors growing close by. As is typical for a solitary tree, the crown gradually spread out broadly in all directions, attaining a relatively spherical shape. In general, branches grow outward and ramify into the space of greater brightness surrounding them. The leaves and branches themselves create darkness so that the outward spreading is toward greater brightness. The crown as a whole is not growing toward the light source (the sun), but toward the brightness of the surrounding atmosphere. In our latitudes, the northern side of a tree will, in more or less subtle ways (leaf size and shape, for example), differ from the southern side, which is exposed to more brightness. Similarly, leaves that are at the outermost edges of the tree differ from leaves situated in the darker interior of the crown. By creating shade, a tree creates an environment for itself, influencing its own growth pattern. A tree is, in part, its own context.
What about the small-crowned white oak with its long, upward-soaring trunk? This specimen grew in the woods. It partnered in growth with red oaks, sugar maples, and red maples. You have to imagine this single tree surrounded on all sides by other trees of similar height. All trees together form one large crown — the forest canopy. By growing up together, perhaps out of an abandoned pasture about 80 to 100 years ago, these trees began growing upward and unfolding. They produced shade for each other, and the dominant growth direction was upward into the light-filled space. The lower branches, which never grew to great size, died off in the increasingly shady environment of the upward-shooting trees. In this way, the long, branchless trunk developed, and we need to imagine the seemingly meager crown of the individual trees as part of the larger, dense, green canopy of the whole forest.
We can now better understand the tree groups in Figure 3. The group consisting of the white ash, American elm, and pignut hickory is a free-standing group in a pasture. The group forms one common crown that resembles that of a single free-standing tree. The common crown of the white oak and the red oak reveals a different context. This pair grows within a relatively bright oak-hickory forest at the edge of a small clearing in moderate light conditions, neither surrounded by brightness from all sides nor illuminated only from above. The common crown form reveals this intermediate situation: the trunks only divide at a considerable height, but then together branch out into a fuller crown than if they had grown in shadier woods.
Directing our gaze toward the form of trees leads us beyond the tree itself. It leads us to a web of relations of which the tree is part. Once you begin to see in such an organic form the tracks of its history and its relations to its surroundings, every meeting with a new tree is a source of excitement, a riddle waiting to be appreciated and deciphered.
Competition, Cooperation — or Neither?
In describing the trees in this article I have consciously avoided the terminology of competition often applied to biological phenomena. Virtually all contemporary ecological and evolutionary studies use competition as the central explanatory framework. On this view, a tree’s genetic propensity is to capture as much light as possible, which maximizes its ability to do photosynthesis, grow, and produce fertile offspring that guarantee the survival of the species. Since every tree has this propensity, and growth creates shade that brings about death, each tree competes with its neighbors to maximize light uptake. Such competition is what brings about the “struggle for existence” that Charles Darwin placed at the heart of his evolutionary theory.
An opposing, much less common interpretation of the trees is that they are cooperating: each tree in a group survives and in so doing does not compete against its neighbors, but rather works together with them, adjusting its growth in relation to the others.

Figure 4. Two different specimens of white oak (Quercus alba). The specimen on the left is a freestanding tree, while the tall, slender tree on the right grew in a forest. (Sketches by C. Holdrege)
Both modes of interpretation are decidedly anthropocentric. Competition and cooperation are concepts drawn from human experience. Shouldn’t we examine critically whether such concepts have any relevance to plants, which are such different creatures from ourselves? Imagine for a moment that we had no first-hand experience as ego-centered agents striving to secure and expand our own existence — the experience that underlies our concept of competition. Would the phenomena of tree growth I have described suggest out of themselves that we are dealing with competing agencies? I don’t think so. Rather, the genetic propensity we ascribe to the plant to maximize light uptake as a survival strategy is a concept we project onto the plant, unaware that we are conceptually infusing its biology with an all-too human psychological characteristic. If we are interested in understanding the trees and not our own reflection in the trees, then we would do well to avoid such an interpretative framework.
Darwin noted that he was using the term “struggle for existence” (what we today call competition) in a “large and metaphorical sense.” He writes in Origin of Species:
A plant on the edge of the desert is said to struggle for life against the drought, though more properly it should be said to be dependent on the moisture. (Darwin 1859)
What a difference between these two ways of expressing the same phenomenon. The first projects competitive agency into the plant. The second simply states in a matter-of-fact way an essential quality of the plant in relation to its environment, namely its reliance on water. I would say, in agreement with Darwin, that the second formulation is undoubtedly more proper because it stays closer to the phenomena themselves and in so doing has the added virtue of leaving one open to further insights that may come with more study of the relations between plant, water, and desert. The idea of competition, by contrast, forces the phenomena into a closed frame shaped by our own, unrelated experience.
To avoid misunderstanding: I am not saying that Darwin or modern ecologists and evolutionary scientists believe plants are intelligent agents scheming to increase the survival of their species. In the barest terms, Darwinian competition is merely the result of the fact that all organisms produce more offspring than can survive. Individual organisms and species have to compete because there is no way all can survive with the limited resources a habitat, an ecosystem, or, in the end, the whole biosphere provides. But even this way of stating the relationships is more interpretive than you might think. It presupposes that you focus on the individual plants or species as things unto them-selves that relate to each other as “others.” As a consequence, species survival is assumed to be writ large in each genome, with competition a necessary outcome and there-fore the dominant mode of species-interaction. If you take this approach far enough, you end up with Richard Dawkins’ “selfish genes,” which become the competing atoms of biology (Dawkins 1990).
Charles Darwin writes,
A plant which annually produces a thousand seeds, of which on average only one comes to maturity, may be more truly said to struggle with the plants of the same and other kinds which already clothe the ground. (Darwin 1859)
If Darwin hadn’t been guided only by the metaphor of competition, he might just as well have emphasized that many seeds and seedlings provide nourishment for birds, soil nematodes, slugs, woodchucks and other creatures. The species does not exist merely “in and of itself;” it is part of a larger whole. This is what we have seen in the development of tree forms. We may be tempted to say that the plants are sacrificing themselves for the greater whole. Do you recognize the strong anthropocentrism? It might be an uplifting thought that touches our feelings to think of self-sacrifice in nature, but it may have little to do with the plants.
To employ concepts such as competition and cooperation is in one way easy and fulfilling. We know these qualities from the inside and we can wield them as a framework within which we place all the phenomena we encounter. If we could use the concept of competition in a free and playful manner, just to see what might show itself through this particular lens, and then shift to another point of view such as the one involving cooperation, we wouldn’t need to worry much about the misuse of these concepts. Their limitations would be counterbalanced by the variety of perspectives, and we would gain through a dialectical process a richer understanding of the world.
But competition is the one reigning perspective used to interpret life. It has become a rarely questioned, unconscious habit of mind. People don’t even notice they are using an interpretative framework and assume that competition is a fact of nature. A concept that is used habitually and unconsciously colors the world we see and limits our under-standing. It no longer illuminates. If we want to cast ever new and fresh light on the nature of things, we need to become much more conscious of the concepts we use and apply them in a discerning and, to use Goethe’s words, “delicate” manner. This effort will help keep science a vital, evolving human enterprise.
References
Buess, Mathias (1992). “The Gobenmatt” in Jochen Bockemühl (ed.), Awakening To Landscape (Dornach, Switzerland.: Naturwissenschaftliche Sektion) pp. 34–55. (This book can be ordered from The Nature Institute.)
Darwin, Charles(1859/1979). The Origin of Species (Middlesex: Penguin Classics) Chapter III, p. 116.
Dawkins, Richard (1990). The Selfish Gene (Oxford: Oxford University Press, second revised edition).
Gleissner, Peter (2005). “Was ist das: ein Typus?” In Volker Harlan (ed.) Wertrund Grenzendes Typusinderbotanischen Morphologie (Nümbrecht: Martina Galunder-Verlag, 2005.)