
What Does it Mean to Be a Sloth?
Craig Holdrege
This article is a lightly edited version of a chapter in Craig's book, Seeing Animals Whole — And Why It Matters, published in 2021 by Lindisfarne Books.
One more defect and they could not have existed.
— George Louis Leclerc, Comte de Buffon
Hence we conceive of the individual animal as a small world, existing for its own sake, by its own means. Every creature is its own reason to be. All its parts have a direct effect on one another, a relationship to one another, thereby constantly renewing the circle of life; thus we are justified in considering every animal physiologically perfect.
— Johann Wolfgang von Goethe
We are losing animals. I do not mean only numerically through the extinction of species. I also mean we are losing them in our understanding. Perhaps it might be better to say we’ve rarely taken animals as whole, integrated beings seriously and therefore they have never really come into view for us. For that reason our scientific and technological culture can so casually manipulate what it does not know. The more we get to know something intimately, the less likely we are to treat it in a purely utilitarian fashion.
Imagine a biotechnologist wondering what causes the sloth to be slow and pondering whether the animal could be mined for “slothful” genes that might be put to therapeutic use in hyperactive children. Or another who wonders whether the sloth might not be a good research model for testing the efficacy of genes from other organisms that enhance metabolic activity. As far as I know, no such research projects are in progress or being planned — and I am glad. But how easily we can come up with ideas that hover in splendid isolation above any deeper concern for the animal itself! We are intrigued and motivated by the seemingly boundless limits of doing the doable. We do not feel limited by ignorance of what we’re dealing with.
This essay is an attempt to show how we can take steps to overcome some of that ignorance — of which we should nonetheless always be mindful — by beginning to grasp something of the organic lawfulness inherent in one animal, the sloth. With all its unique and unusual features, the sloth almost seemed to be prodding me to understand it in an integrated, holistic way. The poet and scientist Goethe set the stage for a sound holistic approach to studying animals, and others have developed his method further.1 I have been influenced and inspired by their work in this study.
The Sloth in its World
Even if you were to look hard and make lots of noise, you would most likely not see the most prevalent tree-dwelling mammal in Central and South America’s rain forests. The monkeys scurry off and perhaps scream. The sloth remains still and hidden.
The rain forest is a tropical ecosystem characterized by constancy of conditions. The length of day and night during the year varies little. On the equator there are twelve hours of daylight and twelve hours of night 365 days a year. The sun rises at 6 am and sets at 6 pm. Afternoon rains fall daily throughout most of the year. The air is humid (over 90%) and warm. The temperature varies little in the course of the year, averaging 25°C (77° F).
Except in the uppermost part of the forest canopy, it is dark in the rain forest. Little light penetrates to the forest floor. The uniformity of light, warmth and moisture — in intensity and rhythm — mark the rain forest. And it is hard to imagine a rain forest dweller that embodies this quality of constancy more than the sloth. From meters below, the sloth is sometimes described as looking like a clump of decomposing leaves or a lichen-covered bough. The sloth’s hair is long and shaggy, yet strangely soft. The fur is brown to tan and quite variable in its mottled pattern. Especially during the wettest times of year, the sloth is tinted green from the algae that thrive on its pelage, which soaks up water like a sponge (Aiello 1985).
Since the sloth moves very slowly and makes few noises, it blends into the crowns of the rain forest trees. It took researchers many years to discover that up to 700 sloths may inhabit one square kilometer of rain forest (Sunquist 1986). Only 70 howler monkeys inhabit the same area.
The sloth spends essentially its whole life in the trees. It hangs from branches by means of its long, sturdy claws, or sits nestled in the forks of tree branches. The contrast to terrestrial mammals in respect to orientation is emphasized by its fur. Instead of having a part on the mid-back, with the hair running towards the belly, as is typical for terrestrial mammals, the sloth’s fur has a part on the mid-belly and the hair runs toward the back.
The sloth moves slowly through the forest canopy — from a few to rarely a few hundred feet in twenty-four hours. On average, sloths were found to move during seven to ten hours of the twenty-four-hour day (Sunquist and Montgomery 1973). The remaining time sloths are asleep or inactive. (Resting is the term often used to describe the sloth’s inactive periods, but this isn’t a sloth-appropriate expression. From what activity is the sloth resting?)
Limbs and Muscles
The sloth’s ability to hang from and cling to branches for hours on end is related to its whole anatomy and physiology. The sloth is about the size of a large domestic cat. It has very long limbs, especially the forelimbs (Figure 1). When hanging, the sloth’s body appears to be almost an appendage to the limbs. Feet and toes are hidden in the fur. Only the long, curved and pointed claws emerge from the fur. The toe bones are not separately movable, being bound together by ligaments, so that the claws form one functional whole, best described as a hook.

Figure 1. The three-toed sloth. (Sketch by Craig Holdrege.)
The two different genera of sloths are named according to the number of claws they possess: the three-toed sloth (Bradypus) has three claws on each limb; the two-toed sloth (Choloepus) has two claws on the forelimb and three on the hind limb. (There are many differences in detail between these two groups of sloths. Most of the specific information referred to in this essay pertains to the three-toed sloth, unless otherwise indicated.)
With its long limbs the sloth can embrace a thick branch or trunk, while the claws dig into the bark. But the sloth can also hang just by its claws on smaller branches, its body suspended in the air. A sloth can cling so tenaciously to a branch that researchers resort to sawing off the branch to bring the creature down from the trees.
All body movements, or the holding of a given posture, are made possible by muscles, which are rooted in the bones. Muscles work by means of contraction. While clinging, for example, some muscles in the limbs — the retractor muscles — are contracted (think of your biceps) while other muscles — the extensor muscles — are relaxed (think of your triceps). When a limb is extended (when the sloth reaches out to a branch) the extensor muscles contract, while the retractor muscles relax. All movement involves a rhythmical interplay between retractor and extensor muscles.
It is revealing that most of a sloth’s skeletal musculature is made up of retractor muscles (Goffart 1971; Mendel 1985a). These are the muscles of the extremities that allow an animal to hold and cling to things and also to pull things toward it. The extensor muscles are smaller and fewer in number. In fact, significant extensor muscles in other mammals are modified in the sloth and serve as retractor muscles. A sloth can thus hold its hanging body for long periods. It can even clasp a vertical trunk with only the hind limbs and lean over backward ninety degrees with freed forelimbs. As the sloth expert M. Goffart points out, “in humans this feat is exceptional enough to be shown in a circus” (Goffart 1971, p.75).
At home as it is in the trees, the sloth is virtually helpless on the ground. Lacking necessary extensor muscles and stability in its joints, a sloth on the ground can hardly support its weight with its limbs. Researchers know little about natural terrestrial movement of sloths. But on rough surfaces captive sloths have been observed slowly crawling (Mendel 1985b). If they are placed on a smooth surface like concrete, their limbs splay to the side. In this position a sloth can only drag its body by finding a hold with the claws of its forelimbs and pulling itself forward, using its strong retractor muscles.
Since the sloth’s main limb movements involve pulling and the limbs do not carry the body weight, it is truly a four-armed and not a four-legged mammal. The hands and feet are essentially a continuation of the long limb bones, ending in the elongated claws and do not develop as independent, agile organs as they do, say, in monkeys. We can also understand the dominance of the retractor muscles from this point of view. The human being, in contrast to most mammals, has arms as well as weight-bearing legs. The arms are dominated by retractor muscles, while the legs have more extensor muscles. Moreover, the arm muscles that move the arm toward the body are stronger than the antagonistic arm muscles that move the arms away from the body. This comparison shows us that the tendency inherent in the arm — the limb that does not carry the body’s weight — permeates the anatomy of the sloth.
A sloth becomes quite agile if the forces of gravity are reduced, as in water. In water a body loses as much weight as the weight of the volume of water it displaces (Archimedes’ Law). The body becomes buoyant, and in the case of the sloth, virtually weightless.
Remarkably, sloths are facile swimmers. . . . They manage to move across water with little apparent effort. Where the forest canopy is interrupted by a river or lake, sloths often paddle to new feeding grounds. With no heavy mass to weigh them down, they float on their buoyant, oversized stomachs. (Sunquist 1986, p. 9)
With its long forelimbs the sloth pulls its way through the water, not speedily, but in a “beautifully easy going” manner (Bullock, quoted in Goffart 1971, p. 94).
On the whole, sloths have little muscle tissue. Muscles make up 40 to 45 percent of the total body weight of most mammals, but only 25 to 30 percent in sloths (Goffart 1971, p. 25). One can understand how the reduction of weight in water allows them to be less encumbered in movement. Sloth muscles also react sluggishly, the fastest muscles contracting four to six times more slowly than comparable ones in a cat. In contrast, however, a sloth can keep its muscles contracted six times longer than a rabbit (Goffart 1971, p. 69). Such anatomical and physiological details reflect the sloth’s whole way of being — steadfastly clinging in a given position, only gradually changing its state.
The tendency to the reduction of muscle tissue can also be found in the head. There is a reduction in the number and complexity of facial muscles (Naples 1985). Through the facial markings the sloth has an expressive face, but this is the expression of a fixed image, rather than expression through movement, since the facial area itself is relatively immobile. The outer ears are tiny and are essentially stationary. The sloth alters the direction of its gaze by moving its head, not its eyeballs. This rather fixed countenance is dissolved at the lips and nostrils, which, as the primary gateways to perceiving and taking in food, are quite mobile.
Paced Metabolism and Fluctuating Body Temperature
Since sloths are externally inactive or asleep a good portion of the twenty-four-hour day and the remaining time is spent slowly moving and feeding, they perform about ten percent of the physiological work of a mammal of similar size (Goffart 1971, p. 59). All metabolic processes are markedly measured in tempo. Sloths use little oxygen, breathe slowly, and the respiratory surface of their lungs is small.
All metabolic activity produces warmth. Warmth is also needed for activity, for example, in the exertion of muscles, which in turn results in more warmth production. Birds and virtually all mammals not only produce warmth, but also maintain a constant body temperature. This is a striking physiological feat. A warm-blooded (endothermic) animal is like a radiating, self-regulating center of warmth. Warmth constantly permeates the whole organism.
Most mammals maintain a constant core body temperature of about 36°C (97°F), which changes very little despite variations in environmental temperatures. For example, in a laboratory experiment a mouse’s internal temperature changes only four tenths of one degree Celsius when the outer temperature rises or falls twelve degrees (Bourlière 1964). Exceptionally, however, a sloth’s body is not so permeated by warmth; in other words, it is not constantly prepared for activity. Its body temperature can vary markedly.
Gene Montgomery and Mel Sunquist, who did extensive field research in Panama on the ecology and physiology of sloths, found that the sloth’s body temperature fluctuated with the ambient temperature (Montgomery and Sunquist 1978). During the morning as the ambient temperature rose, the body temperature also rose. When found on sunny days, sloths were often on an outer branch, belly-side up with limbs extended, basking in the sun. Body temperature usually peaked at about 36-38°C soon after midday. It then began to fall, reaching a low point of about 30-32°C in the early morning. The body temperature was usually about 7-10°C higher than the ambient temperature.
Although sloths are often active at night, their body temperature does not rise in connection with their increased activity. This shows, in contrast to other mammals, that the sloth’s body temperature is less affected by its own activity than by the ambient temperature. According to Brian McNab (1978), the sloth “almost appears to regulate its rate of metabolism by varying body temperature, whereas most endotherms [warm-blooded animals — mammals and birds] regulate body temperature by varying the rate of metabolism.” Raising the outer temperature under experimental conditions is, as Goffart puts it, an effective way of “‘deslothing’ the sloth,” since it then moves around more actively.
A three-toed sloth has an insulating coat of fur comparable to that of an arctic mammal, which seems at first rather absurd for a tropical animal. It has, like an arctic fox, an outer coat of longer, thick hair and an inner coat of short, fine, downy fur. These allow the sloth to retain the little warmth it creates through its metabolic processes. But, characteristically, the sloth cannot actively raise its body temperature by shivering as other mammals do. Shivering involves rapid muscle contractions that produce warmth.
Clearly, the sloth is at home in the womb of the rain forest, which keeps constant conditions like no other terrestrial ecosystem. Not only by virtue of its coloring and inconspicuous movements does the sloth blend into its environment, but through its slowly changing body temperature as well.
Feeding and Orientation
Moving unhurried through the crown of a tree, the sloth feeds on foliage. We usually think of leaf eating (browsing) as an activity done on the ground by mammals, for example, deer. There are, in fact, relatively few leaf-eating mammals in the crowns of trees, although tree leaves are an abundant and constant source of food. Sloths are literally embedded in and surrounded by their food at all times and in all directions. Tropical trees do lose their leaves, but not all at once. Sometimes one and the same tree may lose leaves on one branch, while it sprouts new ones on others.
Sloths don’t eat just any leaves. They seem to prefer younger leaves, and each individual animal has its own particular repertoire of about 40 tree species from which it feeds (Montgomery and Sunquist 1978). A young sloth feeds together with its mother, often licking leaf fragments from the mother’s lips. After its mother departs the juvenile at the age of about six months, the young sloth continues to feed from those species it learned from its mother. This specificity is probably a major factor in the inability to keep three-toed sloths alive in zoos. They usually die of starvation after a short period of time. In contrast, the two-toed sloth is more flexible and survives well in captivity, eating assorted fruits and leaves.
A sloth does not search for leaves with its eyes. Its eyesight is very poor and it is short-sighted (Goffart 1971, pp. 106ff.; Mendel et al. 1985). The eyes lack the tiny muscles that change the form of the lens to accommodate for changing distances of objects. As if to emphasize the unimportance of its eyes, the sloth can retract them into the eye sockets. The pupils are usually tiny, even at night. Clearly, a sloth does not actively penetrate its broader environment with its vision, as do most arboreal mammals like monkeys.
Sight and hearing (the latter also not very developed in sloths) are the two senses through which animals perceive and react to stimuli at a distance. The sloth makes little use of these senses, relying much more on a sense that entails drawing the environment into itself: the sense of smell.
I placed a sloth, hungry and not too disturbed, on an open area under the bamboos, and planted four shoots twenty feet away in the four directions of the compass. One of these was Cecropia [a primary food of three-toed sloths] camouflaged with thin cheesecloth, so that the best of eyesight would never identify it, and placed to the south, so that any direct wind from the east would not bring the odor too easily. The sloth lifted itself and looked blinkingly around. The bamboos thirty feet above, silhouetted against the sky, caught its eye, and it pitifully stretched up an arm, as a child will reach for the moon. It then sniffed with outstretched head and neck, and painfully began its hooking progress toward the Cecropia. . . . Not only is each food leaf tested with the nostrils, but each branch. . . . (Beebe 1926, p. 23)
So we should not imagine a sloth looking at its food. Rather, a sloth immerses and orients itself in a sea of wafting scents.
When the sloth is in the immediate proximity of leaves it feeds on, it will hook the branch with the claws of a fore- or hind limb and bring the leaves to its mouth. Having no front teeth (incisors), it tears off the leaves with its tough lips. It chews the leaves with its rear, peg-like teeth. Unlike most leaf-eating mammals (for example, deer), the sloth lacks many deeply rooted, hard, enamel-covered grinding teeth. The sloth also has comparatively few teeth (18 compared to 32 in most deer). Moreover, the teeth lack enamel altogether and wear easily. In compensation, the teeth grow slowly throughout the animal’s life. There is no change of teeth from milk to permanent dentition. Growth and wear are in balance.
While feeding, the sloth is continuously chewing and simultaneously moving food backward with its large tongue in order to swallow. Sloths can feed in all positions, even hanging upside down. A young, captive two-toed sloth showed “decided preference for eating upside down in the manner of adult sloths at eight months” (Goffart 1971, p. 114).

Figure 2. The three-toed sloth. Note the orientation of the head. (Sketch by Craig Holdrege.)
The sloth can move its head in all directions, having an extremely flexible neck. Imagine a sloth hanging from all four legs on a horizontal branch. In this position the head looks upward (like when we lie in a hammock). Now the sloth can turn its head — without moving the body — 180 degrees to the side and have its face oriented downwards. As if this were not enough, the sloth can then move its head vertically and face forward — an upright head on an upside down body (Figure 2)! When it sleeps, a sloth can rest its head on its chest.
The sloth’s neck is not only unique in its flexibility, but also in its anatomy. Mammals have seven neck (cervical) vertebrae. The long-necked giraffe and the seemingly neckless dolphin — to mention the extremes — both have seven cervical vertebrae. This fixed mammalian pattern is abandoned by only the sloth and the manatee. The three-toed sloth usually has nine and the two-toed sloth has between six and nine cervical vertebrae.
Centered in its Stomach
Digestion in the sloth occurs at an incredibly slow rate. In captive animals “after three or six days of fasting the stomach is found to be only slightly less full” (Britton 1941). Leaves are hard to digest and not very nutrient-rich, consisting primarily of cellulose and water. Only with the help of microorganisms in the stomach can the sloth digest cellulose, breaking it down into substances (fatty acids) that can be taken up by the blood stream.
The sloth’s stomach is four-chambered like those of ruminants (cows, deer, and so on) and is clearly the center of the digestive process. The stomach is enormous relative to the animal’s overall size. It takes up most of the space of the abdominal cavity and, including contents, makes up 20 to 30 percent of total body weight. Nonetheless, digestion takes a long time. On the basis of field experiments, Montgomery and Sunquist (1978) estimate that it takes food about ten times longer to pass through a sloth than through a cow. Moreover, the sloth also digests less of the plant material than most other herbivores.
Through its stomach a mammal senses hunger. Most grazing mammals spend a large part of their time eating, so that food is continuously passing through their digestive tract. The sloth is, once again, an atypical herbivore since it feeds for a comparatively small portion of its day. A small rain forest deer, the same size as a sloth, ate six times as much during the same period of time (Beebe 1926). The howler monkey, which also lives in the canopies of Central and South American rain forests and whose diet comprises only about 50% leaves, eats about seven times as many leaves as do sloths. With its slow metabolism and digestion, the sloth’s stomach remains full, although the animal eats so little.
As a stark contrast, we can think of carnivores like wolves or lions that regularly, as a normal part of their lives, experience empty stomachs. Their hunting drives are directly connected to their hunger. Hunger brings about the maximal aggressive activity of these animals. When a lion has gorged itself on forty pounds of meat, it becomes lethargic and sleeps for an extended period. The sloth’s constantly full stomach is a fitting image for its consistently slow-paced life as well as, it seems, a physiological condition for it: “starvation makes [sloths] hyperactive” (Goffart 1971 p. 113).
After about a week of feeding, sleeping and external inactivity, a change occurs in the sloth’s life. It begins to descend from its tree. Having reached the forest floor, it makes a hole in the leaf litter with its stubby little tail. It then urinates and defecates, covers the hole, and ascends back into the canopy, leaving its natural fertilizer behind. (The two-toed sloth has no tail and leaves its feces lying on the leaf litter.)
The feces, the product of sloth metabolism, decompose very slowly. The hard pellets can be found only slightly decomposed six months after defecation. Normally, organic material decomposes rapidly in the warm and moist conditions of the rain forest. For example, leaves decompose within one or two months (a process that can take a few years in a temperate-climate forest). Ecologically, sloth excrement “stands out as a long-term, stable source [of nutrients] . . . and may be related to stabilizing some components of the forest system. . . . Sloths slow the normally high recycling rates for certain trees . . .” (Montgomery and Sunquist 1975, p. 94). Sloths contribute not only slow movement to the rain forest but slow decomposition as well!
It is estimated that a sloth can lose up to two pounds while defecating and urinating, more than one fourth of its total body weight (Goffart 1971, p. 124). If one imagines a sloth with a full stomach (which it always seems to have) just prior to excreting, then more than half of its body weight is made up of its food, waste and digestive organs! This quantitative consideration points to the qualitative center of gravity in the animal’s life. But the sloth’s stomach is more like a vessel that needs to remain full than a place of intensive muscular activity, secretion, mixing and breaking down, as it is in the cow, for example.
Stretching Time
The sloth researcher William Beebe wrote in 1926: “Sloths have no right to be living on this earth, but they would be fitting inhabitants of Mars, whose year is over six hundred days long.” Beebe was deeply impressed by the way in which sloths “stretch” time, another way of characterizing their slowness. We have seen how this quality permeates every fiber of their day-to-day existence. It is therefore not so surprising to find that the development of sloths takes a long time.
Sloths have a gestation period of four to six months, compared to a little over two months in the similar-sized cat. One two-toed sloth in a zoo gave birth after eight-and-one-half months. Initially more surprising was the rediscovery of a female sloth in the rain forest 15 years after she had been tagged as an adult. This means she was at least 17 years old, “an unusually long life span for such a small mammal” (Montgomery, quoted in Sunquist 1986). Thus, regarding time, the qualities of the sloth certainly speak a unified language.
Gravity and the Skeleton
If we look for the embodiment of fixed form in the organ systems of a mammal, then we come to the skeleton. The bony skeleton gives the mammal its basic form and is the solid anchor for all movement. The limb bones develop their final form in relation to both gravity and their own usage. An injured quadruped mammal will lose bone substance in the leg it is not using, which does not carry any weight. Conversely, in the other three limbs bone matter is laid down to compensate for the increase in weight carried and muscular stress.
The sloth has a special relation to gravity. As mentioned earlier, the limbs hold the hanging body; they do not carry it (Figure 3). The sloth gives itself over to gravity rather than resisting it and living actively within it via its skeletal system. A sloth kept on the ground in a box developed raw feet from the unaccustomed pressure (Beebe 1926).
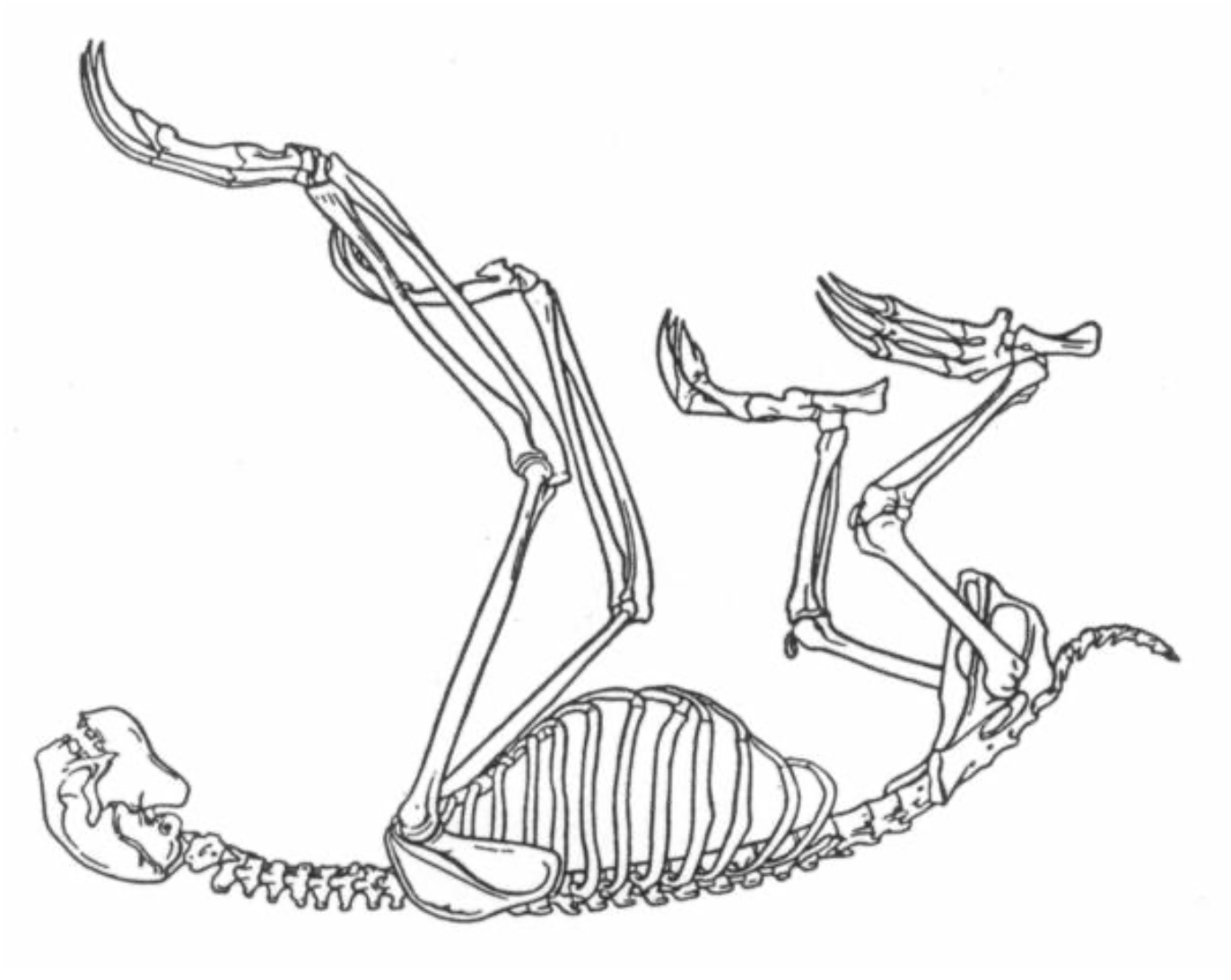
Figure 3. Skeleton of a three-toed sloth. (Reprinted from Young 1973, 600.)

Figure 4. Skeleton of a horse. (Reprinted from Tank 1984, 108.)
The other pole in relation to gravity is represented by hoofed mammals like deer, horses or giraffes. By virtue of their skeletal architecture they can relax their muscles and even sleep while standing. Their legs are solid, stable columns that carry the body’s weight (Figure 4). In contrast, the sloth has very loose limb joints. In his detailed study of the limbs of the two-toed sloth, Frank Mendel (1985a, p.159) points out how unusual the “poorly reinforced and extremely lax joint capsules” are. This anatomical peculiarity allows a wide range of limb movement and is connected with the fact that the joints are not subject to compression as they are in weight-bearing limbs. Through clinging and hanging, the joints of a sloth are being continually stretched. Similarly, the sloth has a flexible, curved spine. The hoofed mammal, in contrast, has a stiff, straight spine, from which the rib cage and internal organs of the torso are suspended. A deer would be as ungainly in a tree as a sloth is on the ground.
This contrast is mirrored in the teeth. Hoofed mammals have deeply rooted, very hard teeth with ridges of enamel that withstand the toughness of grass. Enamel is the hardest substance a mammal can produce, and, as already mentioned, sloth teeth have no enamel coating. In addition, more than in other mammals, the form and chewing surfaces of the sloths’ teeth are sculptured during usage. “Since sloth teeth acquire their individual characteristics through wear, it is very difficult to distinguish the young of one genus from those of another based upon shape or location of dentition” (Naples 1982 p. 18). In other mammals — especially the grazers — the teeth are preformed with all their crown cusps and ridges before they erupt. The sloth’s teeth emerge as simple cones and take on a characteristic form in the course of life.
The sloth is, in this sense, formed from the outside. In a related way we see this tendency in its coloring, which arises not only from hair pigmentation but also through algae from the surroundings. Similarly, its temperature varies with the ambient temperature.
From a different vantage point we can say: incorporating solidity and stability into the skeleton allows a quadruped mammal to live actively within gravitational forces. In giving itself over to gravity, the sloth incorporates inertia. We see inertia in its movements and digestion. The sloth is a bit like the clump of leaves or the alga-covered tree trunk it outwardly resembles.
Drawing In
Active arboreal mammals, like monkeys, have, of course, nothing of the skeletal rigidity of ground-dwelling quadrupeds. They have flexible joints and muscular agility that allow for actively swinging, jumping, and grasping. A sloth lacks the quick and nimble dexterity of monkeys, although it possesses a flexible spinal column (especially in the neck region) and limber fore- and hind limbs. A sloth can twist its forelimb in all directions and roll itself into a ball by flexing its vertebral column.
Characteristically, the sloth makes use of this flexibility for its slow movements while feeding and also for protecting itself from a predator by curling up into a ball. The monkey, in contrast, engages in light and springy movements. This leads us to a slightly different way of characterizing the sloth. A primary gesture is that of pulling in or retracting. It doesn’t project actively out into its surroundings.
We can see this tendency in the head. The head is the center of the primary sense organs through which an animal relates to its environment. As we have seen, the eyes and ears are not the sloth’s main senses. The outer ears (pinnae) are tiny and hardly visible on the head and the eyes can retract in their sockets. Both of these characteristics reveal externally the muted function of these organs within the whole animal. They also let the head appear as a broadened neck. But this appearance also has a deeper anatomical basis, since the first cervical vertebra (the so-called atlas) is nearly as wide as the widest part of the skull.

Figure 5. Skulls of a three-toed sloth (top, left), old world monkey (top, right), and horse (bottom). (Drawings by Craig Holdrege; the skulls are not drawn to scale.)
The skull itself is rounded and self-contained — superficially resembling a monkey’s skull more than a grazing herbivore’s (Figure 5). Most herbivores have an elongated snout that they use as a limb — standing as they do on all four legs — to reach their food. The sloth’s forelimbs have this function and thus its snout is short. The premaxillary bones — important in forming the elongate mammalian snout — are tiny in the sloth. Moreover, the upper jawbones (maxillae) and the nasal bones are also short in the sloth. The sloth’s skull does not project forward.
We have seen that the sense of smell is the sloth’s primary sense and that its gesture is to draw in, in contrast to the more outwardly projecting senses of sight and hearing. When we see these facts together with the others, such as the dominance of retractor muscles, then the sloth’s special orientation to its surroundings comes more clearly into view.
The Sloth as a Habitat
As if to emphasize its passive, somewhat withdrawn character, the sloth functions as a habitat for myriad organisms. I have mentioned the algae that live in its fur, giving the pelage a greenish tinge. In addition to the usual ticks and flies that infest the skin and fur of other mammals, a number of sloth-specific moth, beetle, and mite species live on the sloth and are dependent upon it for their development. The sloth moths and beetles live as adults in the sloth’s fur. Some species live on the surface and others inhabit the deeper regions of the fur. They are evidently not parasitic; their source of food is unknown.
When the sloth descends from a tree to defecate and urinate, female moths and beetles fly off the animal and lay their eggs in the sloth’s dung. The wings of one moth species break off soon after they inhabit the sloth, so that they are incapable of flying. Consequently they must crawl off the sloth to reach the dung. The sloth’s relatively long period of defecation, which lasts a few minutes, gives the insects the time they need. In this way the slowness of the sloth serves these most “slothful” of sloth moths!
The larvae develop in and feed on the dung (which, you remember, decomposes slowly). The larvae pupate in the dung and the winged adult moths (or beetles) fly off to inhabit another sloth. Various species of insects and mites inhabit any given sloth, and the numbers of specimens of each species varies greatly, ranging from a few to over a hundred.
The sloth has been observed grooming its fur. This is typical mammalian behavior and does rid an animal of some of its “pests.” From this utilitarian point of view, the sloth’s grooming is not very effective. Typically, sloths groom slowly, and sloth moths “may be seen to advance in a wave in front of the moving claws of the forefoot, disturbed, but by no means dislodged from the host” (Waage and Best 1985, p. 308). Clearly, the measured pace of life, the unique excretory habits, and the consistency of dung allow the sloth to be a unique habitat for such a variety of organisms.
Sensing a Boundary
The expression of pain is a barometer for the way an animal experiences its own body in relation to the environment. Pain is one way an animal experiences the external world penetrating and harming its biological integrity. Here’s an example from a family that kept a sloth at their home in Brazil:
‘Sloth burning!’ . . . we leap to our feet and run frantically round trying to discover where [the sloth] has fallen asleep. On the kitchen stove? No! On the water heater in the bathroom? No! There he is on top of the floor lamp in the drawing-room, with his bottom touching the big electric bulb! . . . We struggle to get him down, but he clings desperately to his perch, refusing to budge and protesting with many ah-eees against our unwarranted disturbance of his slumbers. (Tirler 1966, p. 27)
Sloths are reported to “survive injuries that would be deadly within a short time to other mammals” (Grzimek 1975). “I have known a sloth to act normally for a long time after it had received a wound which practically destroyed the heart . . .” (Beebe 1926, p. 32). These examples show that the sloth does not seem to notice an intrusion of its boundaries and continues to live despite them. Its body is not imbued with sensitive reactive presence.
A Further Dimension of Wholeness: The Environment?
Where does the sloth end? This seemingly naïve question points to a problem and, at the same time, to a task. The problem is the way we think of an organism in relation to its environment. The environment is that with which an animal interacts. Inasmuch as the sloth eats leaves, leaves belong to its environment. In the moment it is interacted with (for example, in feeding, smelling, moving), the environment is part of the animal. We could also say, the animal is part of its environment. The environment as a functional concept is inseparable from the organism (Riegner 1993). The corpus of an animal with its definite outline — what we call the body — fills a definable volume in space. But the animal’s activity carries beyond this corpus. And the environment is part of this activity; without the environment there would be no activity.
It may seem strange to say that the environment is not outside the animal. But this is only because we use spatial terms to describe something functional. Because it is more natural for us to think about the world in the framework of objects, we consider the organism here and the environment there. But this accounts only for the bodily aspect of the organism, and not its functional and behavioral relations. When we shift our focus from the body as a thing to the body as focal point of activity, then the organism encompasses, firstly, all activities radiating to and from this focal point and, secondly, everything we consider to be outside the organism before we change to the functional mode of viewing — leaves, branches, scents, and so on. (I have spoken and will continue to speak of organisms and their environments, otherwise I would have to create some new, probably cumbersome, terminology. There is no problem using existing terminology as long as we can see through it to the expanded concept.)
Viewed in this way, organisms actually interpenetrate. Sloth, tree, sloth moth, and algae are all part of each other. We can, therefore, in principle, understand how an ecological community, an ecosystem, and even the whole earth can be considered as further dimensions of organisms. Speaking of the earth as an organism is then no longer merely an analogy, but becomes a reality one has in part begun to grasp — in this case, through the sloth. (And because we have already seen a part, we have also caught a glimpse, in it, of the whole!) It remains the task of a truly holistic or organismic ecology to concretely apply this way of viewing to ecological phenomena.
Is There a Cause of Slothfulness?
In his compendium on sloths (1971), M. Goffart includes one section entitled “Slothfulness.” He describes observations in the field, experimental results, and the hypotheses of scientists concerning the causes of slothfulness. Various possible explanations are brought forth: small heart, slow speed of muscle contraction, low body temperature, low rate of thyroid function, and so forth. He describes the shortcomings of each particular hypothesis and concludes that the “evidence as to the real causes of slothfulness is thus far from complete” (p. 95). If he were writing today, he might include conjectures about genetic mechanisms.
Goffart points out, for example, that the sluggish koala has a constant body temperature of 36 degrees Celsius. Since this is a normal body temperature for mammals, it seems evident that it cannot be causing the koala’s sluggishness. Since causes are assumed to be general, he concludes that temperature will also not be the cause of slothfulness in sloths.
Goffart assumes that the causes of slothfulness will one day be found; we are just lacking the necessary information. I question this assumption and believe that such an example shows, in fact, primarily the limitations of the conceptual framework. In treating aspects of an organism as potential causes, we conceptually lift them out of the organism. Then we think of them affecting things in the organism as though they were not part of it. By so doing we can think in general terms of the factor “body temperature” as a cause, as if separate from the organism.
But every time we carry through this process we realize that our conceptual scheme doesn’t fit reality, because we are confronted with mutual relations, all of which express something of the animal as a whole. If we drop this scheme, then it becomes interesting that body temperature evidently means two very different things in the koala and the sloth. Instead of looking for genetic or physiological causes that we assume have general validity, we look at the unique expression of physiological facts in the given context. We take the unique integrity of each animal seriously.
It is second nature for a scientist to inquire after the causes of what is under investigation. Some would even say this is the task of science. But in the context of organisms this method alone is not adequate. Putting it a bit radically, biologists would do well to eradicate the term “cause” from their vocabulary and use the more modest and open term “condition.” What genetic, physiological, behavioral, and ecological studies can show is how aspects of an organism provide mutual and changing conditions for each other. This knowledge is extremely valuable as long as we don’t separate it from the organism as a whole. In fact, it can be the gateway to understanding the organism as an integrated whole.
Encircling the Unspeakable: The Animal as a Whole
I’d like to return to the statements quoted at the beginning of this essay: George Louis Leclerc, Comte de Buffon was a well-known 18th-century French scientist. He studied many animals, among them the sloth, about which he said: “one more defect and they could not have existed” (quoted in Beebe 1926). He considered the sloth’s remarkable characteristics to be defects. And they are, if you take the point of view of a horse, eagle, jaguar, or human being. But as naturalist William Beebe countered, “a sloth in Paris would doubtless fulfill the prophecy of the French scientist, but on the other hand, Buffon clinging upside down to a branch of a tree in the jungle would expire even sooner” (Beebe 1926 p. 13).
Buffon takes a standpoint outside the animal. I have followed Goethe’s suggestion and tried to view the sloth on its own turf. He wrote:
Hence we conceive of the individual animal as a small world, existing for its own sake, by its own means. Every creature is its own reason to be. All its parts have a direct effect on one another, a relationship to one another, thereby constantly renewing the circle of life; thus we are justified in considering every animal physiologically perfect. (Goethe 1995, p. 121)
I have made use of comparison, but not to describe what the sloth “should” have in order to be a reasonable animal. The animals described by way of comparison shed light on the sloth, allowing its uniqueness to stand out all the more perceptibly. When Goethe calls an animal “perfect,” he means that each animal has its own unique way of being — its specific integrity that we can try to understand. But this is no simple matter. Goethe recognized that “to express the being of a thing is a fruitless undertaking. We perceive effects and a complete natural history of these effects at best encircles the being of a thing. We labor in vein to describe a person’s character, but when we draw together actions and deeds, a picture of character will emerge” (1995, p. 121; translation modified by CH). In trying to paint a picture of the sloth, I have discussed many details, because through them the whole lights up. Henri Bortoft puts it well when he says, “The way to the whole is into and through the parts. The whole is nowhere to be encountered except in the midst of the parts” (1996, p. 12).
This emergent picture of the whole does not and cannot encompass the totality of its characteristics. One can always discover new details. I am not striving for totality, but rather for wholeness. Our understanding hinges on our ability to overcome the isolation of separate facts and to begin to fathom the animal as a whole, integrated organism. The whole is elusive, and yet, at every moment, potentially standing before the mind’s eye. When we begin to see how all the facets of the animal are related to each other, then it comes alive for us. Or, putting it a bit differently, the animal begins to express something of its life in us. Every detail can begin to speak “sloth,” not as a name, but as a qualitative concept that a definition can do little justice to.
I have tried to describe the sloth in a way that allows us to catch glimpses of its wholeness. I can now refer to such characteristics as slowness, inertia, blending in with the environment, receding or pulling in and not actively projecting outward. Each expression is a different way of pointing to the same coherent whole. Taken alone, as abstract concepts or definitions, they are empty. They are real only inasmuch as they light up within the description or perception of the animal’s characteristics. But they are not things like a bone or an eye. They are, in context, vibrant concepts that reveal the animal’s unique way of being.
Let’s return to the sloth, high in the crown of a rain-forest tree, hanging from or nestled on a branch. In its outer aspect, it blends in with its environment. There are no sudden or loud movements. The sloth’s green-tinged, mottled brown coat lets it optically recede into the wood and foliage of its surroundings. And like the tree bark, the sloth’s fur is teeming with insect life. The sloth’s body temperature rises and sinks with the ambient temperature.
The round form of its head is the anatomical image of the way in which the sloth does not actively project into its environment. There are no large, movable, reactive outer ears. and the eyes are rarely, if ever, moved. The sloth has no protruding snout. It draws the scents of the environment, especially of the leaves it feeds upon, into its nose. But much of the day the sloth is curled up, unaware of the world around it. Even when awake, the sloth seems not to live as intensely in its body as other mammals, being quite insensitive to pain.
The sloth does not carry its own weight; rather, it clings to an outer support. Its skeletal system is not characterized by stability, but by looseness. This laxity allows the sloth to adopt positions that would be contortions in other animals. The sloth makes mostly steady pulling movements with its long limbs, a capacity based on the dominance of retractor muscles.
The sloth develops slowly in the womb and has a long, slow life. It moves slowly through the crowns, feeding on the leaves that surround it from all sides, bathing, as it were, in its food source. The leaves pass through the animal at an almost imperceptibly slow rate. The sloth’s stomach is always filled with partially digested leaves. Even its dung disappears slowly, despite the warm and humid rain forest climate that normally accelerates decomposition processes.
The sloth brings slowness into the world.
~ ~ ~
This is a revised version of an article originally published in the Newsletter of the Society for the Evolution of Science vol. 14, no. 1 (1998), pp. 1-26.

If you enjoyed this article, a collection of other whole-organism profiles can be found in Craig Holdrege’s latest book, Seeing The Animal Whole - And Why It Matters (Lindisfarne Books, 2021), available in our bookstore.
Notes
For general expositions of Goethe’s method see: Bortoft 1996; Goethe 1995; Steiner 1988. For the biological application of a holistic methodology see: Portmann 1967; Schad 1977; Schad, ed., 1983; Riegner 1993 and 1998; Kranich 1995 and 1999; Suchantke 2001 and 2002; Holdrege 1998, 2004, and 2005.
References
Aiello, Annette (1985). “Sloth Hair: Unanswered Questions,” in The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas, edited by G. Gene Montgomery, pp. 213-218. Washington, D.C.: Smithsonian Institution Press.
Beebe, William (1926). “The Three-toed Sloth,” Zoologica vol. 7, pp. 1-67.
Bortoft, Henri (1996). The Wholeness of Nature: Goethe’s Way toward a Science of Conscious Participation in Nature. Hudson NY: Lindisfarne Press.
Bourlière, Francois (1964). The Natural History of Mammals. New York: Alfred A. Knopf, pp. 252 ff.
Britton, W. S. (1941). “Form and Function in the Sloth,” Quarterly Review of Biology vol. 16, pp. 13-43, 190-207.
Goethe, Johann W. von (1995). Goethe: Scientific Studies (Collected Works, vol. 12, ed. Douglas Miller). Princeton: Princeton U. Press.
Grzimek, Bernhard (1975). Grzimek’s Animal Life Encyclopedia, vol. 11 (Mammals II). New York: Van Nostrand Reinhold.
Holdrege, Craig (1998). “Seeing the Animal Whole: The Example of Horse and Lion,” in Seamon and Zajonc 1998, pp. 213-32.
Holdrege Craig (2004). The Flexible Giant: Seeing the Elephant Whole. Ghent NY: The Nature Institute.
Holdrege Craig (2005). The Giraffe’s Long Neck: From Evolutionary Fable to Whole Organism. Ghent NY: The Nature Institute.
Kranich, Ernst-Michael (1995). Wesensbilder der Tiere. Stuttgart: Verlag Freies Geistesleben.
Kranich, Ernst-Michael (1999). Thinking Beyond Darwin. Great Barrington MA: Lindisfarne Press.
Montgomery, G. G. and M. E. Sunquist (1975). “Impact of Sloths on Neotropical Forest Energy Flow and Nutrient Cycling,” in Tropical Ecological Systems, edited by Frank.
B. Golley and Ernesto Medina. New York: Springer Verlag, pp. 69-98.
Goffart, M. (1971). Form and Function in the Sloth. Oxford and New York: Pergamon Press.
McNab, Brian K. (1978). “Energetics of Arboreal Folivores: Physiological Problems and Ecological Consequences of Feeding on an Ubiquitous Food Supply,” in The Ecology of Arboreal Folivores, edited by G. Gene Montgomery. Washington, D.C.: Smithsonian Institution Press, pp. 153-62.
Mendel, Frank (1985). “Adaptations for Suspensory Behavior in the Limbs of Two-toed Sloths,” in The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas, edited by G. Gene Montgomery. Washington, D.C.: Smithsonian Institution Press, pp. 151-62.
Mendel, Frank (1985b). “Use of hands and Feet of Three-toed Sloths (Bradypus variegatus) during Climbing and Terrestrial Locomotion,” Journal of Mammalogy vol. 66, pp. 359-66.
Mendel, Frank, David Piggins and Dale Fish (1985c). “Vision of Two-toed Sloths (Choloepus),” Journal of Mammalogy vol. 66, pp. 197-200.
Montgomery G. Gene and M. E. Sunquist (1978). “Habitat Selection and Use by Two-toed and Three-toed Sloths,” The Ecology of Arboreal Folivores, edited by G. Gene Montgomery. Washington, D.C.: Smithsonian Institution Press, pp. 329-59.
Naples, Virginia L. (1982). “Cranial Osteology and Function in the Tree Sloths, Bradypus and Choloepus,” American Museum Novitates vol. 2739, pp. 1-41.
Naples, Virginia L. (1985). “The Superficial Facial Musculature in Sloths and Vermilinguas (Anteaters),” in The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas, edited by G. Gene Montgomery. Washington, D.C.: Smithsonian Institution Press, pp. 173-89.
Portmann, Adolf (1967). Animal Forms and Patterns. New York: Schocken Books.
Riegner, Mark (1993). “Toward a Holistic Understanding of Place: Reading a Landscape through Its Flora and Fauna,” in Dwelling, Seeing and Designing: Toward a Phenomenological Ecology, edited by David Seamon. Albany NY: SUNY Press.
Riegner, Mark (1998). “Horns, Hooves, Spots, and Stripes: Form and Pattern in Mammals,” in Seamon and Zajonc 1998, pp. 177-212.
Schad, Wolfgang (1977). Man and Mammals: Toward a Biology of Form. Garden City NY: Waldorf Press.
Schad, Wolfgang (ed.) (1983). Goetheanistische Naturwissenschaft vol. 3: Zoologie. Stuttgart: Verlag Freies Geistesleben.
Seamon, David and Arthur Zajonc (eds.) (1998). Goethe’s Way of Science: Toward a Phenomenology of Nature. Albany NY: SUNY Press.
Steiner, Rudolf (1988). Goethean Science. Spring Valley NY: Mercury Press.
Suchantke, Andreas (2001). Eco-Geography. Great Barrington MA: Lindisfarne.
Suchantke, Andreas (2002). Metamorphose. Stuttgart, Germany: Verlag Freies Geistesleben.
Sunquist, Fiona (1986). “Secret Energy of the Sloth,” International Wildlife vol. 16, pp. 6-10.
Sunquist, M. E. and G. G. Montgomery (1973). “Activity Patterns and Rates of Movement of Two-toed and Three-toed Sloths (Choloepus hoffmanni and Bradypus infuscatus),” Journal of Mammalogy vol. 54, pp. 946-54.
Tank. W. (1984). Tieranatomie für Künstler. Ravensburg, Germany: Otto Maier Verlag.
Tirler, Hermann (1966). A Sloth in the Family. London: The Harvill Press.
Waage, J. K. and R. C. Best (1985). “Arthropod Associates of Sloths,” in The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas, edited by G. Gene Montgomery. Washington, D.C.: Smithsonian Institution Press, pp. 297-322.